
Vajon hogyan jött létre az élet? Miféle magból nőhetett ki az élet fája? Hogyan indult el ez a folyamat? Ezen kérdésekre próbálok választ adni az „Ötödik csoda rövid történetében”.
Napjainkban eléggé sokszínű a biológiai élet. A szénalapú organizmusok benépesítették a bolygó minden zugát szárazföldön, vízen és a levegőben egyaránt. Azonban ne gondoljuk, hogy a sokszínű élet öröktől fogva létezett, hiszen a Föld születése után sok millió évig nem alakult ki komplex élet, de még DNS, RNS sem. A kietlen és zord földi környezetet egy apró szintetizáló baktérium tette élhetővé. Előbb megtisztította a légkört és a tengerek tömegét, amelyeket ez után megtöltött oxigénnel. Ebből az új és erős energiaforrásból tört elő az élet. Napjainkban eléggé sokszínű a biológiai élet. A szénalapú organizmusok benépesítették a bolygó minden zugát szárazföldön, vízen és a levegőben egyaránt. Azonban ne gondoljuk, hogy a sokszínű élet öröktől fogva létezett, hiszen a Föld születése után sok millió évig nem alakult ki komplex élet, de még DNS-, RNS sem. A kietlen és zord földi környezetet egy apró szintetizáló baktérium tette élhetővé.
Az élet eredetének kérdése két és fél évezrede foglalkoztatja a filozófusokat, a teológusokat és tudósokat egyaránt. Az evolúció és természetes kiválasztódás óta csak két átfogó elmélet alakult ki az élet kialakulásáról. A kettő közül az egyik a földi vagy terresztrikus, míg a másik a földönkívüli, ún. extraterresztrikus kialakulás helyezi előtérbe.
A földi preferenciákat szorgalmazók szerint az élet a Földön kémiai kotyvasztással alakult ki az „őslevesben”. A másik szerint – legfőképpen Hoyle (1972) és Wickramasinghe szerint – az élet csírái az űrből érkeztek, életképes mikrobák formájában. Ezt nevezik pánspermia-hipotézisnek. A földi viszonyokat figyelembe véve kialakult egy harmadik elképzelés is – egyben Davies (2000) elképzelése is – miszerint az élet a Föld belsejében született, kilométerekre a szilárd kéreg alatt, ahol a geotermikus hő katlanszerű körülményeket hozott létre, amelyhez párosult a felszín alatti zóna vegyi potenciálja. Jelen esetben szorítkozzunk arra, hogy az élet a Földön alakult ki. Az élet kialakulásáról explicit bizonyítékunk nem lehetnek, pusztán rejtett, közvetett implicit bizonyítékaink lehetnek, egy kutatólaboratóriumban, vagy az óceánokban. Ezek közül kerül bemutatásra négy elmélet.
Az ősleves
Sokáig úgy gondolták, hogy a 20. század közepén „kotyvasztott” őslevesben alakult ki az élet. MILLER és UREY (1953) kutatópáros 1953-ban egy lombikban vizet és a korai Föld feltételezett gázait (szén-dioxid, nitrogén, metán, ammónia, hidrogén) keverte össze. A keveréket villámlást utánozó elektromos kisülésekkel stimulálta. A kutatópáros néhány hónap múlva megvizsgálta a lombikot, és meglepődve tapasztalta, hogy az őslevesben szerves molekulákat, köztük néhány aminosavat figyelt meg. Ezt a megfigyelést megalapozva sokáig az a hit élt, hogy az élővilág egy „őslevesben” fogant meg. Azonban idővel kiderült, hogy a Föld ősi légköre sosem volt metánban, ammóniában és hidrogénben gazdag. Ebből következően az ősatmoszféra szimulációja csalódást keltett, ugyanis a „megmaradt” szén-dioxid, nitrogén és nyomokban metán elegyéből elektromos kisüléssel bombázva alig ha lesz szerves anyag, csupán csekély aminosav keletkezett. Egyéb sebekből is vérzett az „ősleves” elmélet, hiszen termodinamikailag nem ésszerű. A molekuláris biológia termodinamikai tétele szerint, a molekulás elektronokat akarnak veszíteni, vagy felvenni; az ellentétes töltést vonzzák; az azonosat taszítják, vagy a hasonló jellegű molekulákkal élnek együtt. Ebből következően az „őslevesben” nincs semmi, ami reagálni akarna, legalábbis úgy nem, mint az oxigén és a hidrogén. Mivel a megmaradt gázok (szén-dioxid, nitrogén, metán) nem teremtett egyensúlytalansági helyzetet, így nem alakulhatott ki semmi, legkevésbé sem DNS és RNS.
A szilikát alapú élet
Amikor az ősleves elmélet elkezdett sebeiből vérezni, egy skót kémikus, CARINS-SMITH (1982, 1985) felvetette a szervetlen ásvány elméletét. Hipotézise szerint 3 milliárd évvel ezelőtt jött létre a DNS, amelyet egy másik entitás előzött meg, a szilíciumé. Szerinte az élet eredete önmagukat másoló szervetlen kristályokon, például szilikátokén alapult. Így az eredeti replikátorok – amelyek a DNS-t kódolják – szervetlen anyagok kristályai voltak. Cains-Smith szerint szerves molekulák kiemelkedő szerepet játszottak a szervetlen kristály replikátorainak nem másoló eszközei között. Ilyen például a montmorillonit, amely feldarabolja a karboxi-metil-cellúzt, viszont kis mennyiségben a karboxi-metil-cellúz összetartja a montmorillontitot (DAWKINS 2011). Elmélete szerint az RNS-t eleinte tisztán szerkezeti célokra használták a kristályok, csak jóval később foglalta el valódi szerepét az RNS, és 3 milliárd év múlva a DNS.
A bolondok aranya
A ’70-es évek elején a Galapágos-hasadéknál melegvíz feltörést tapasztaltak, amelyet az Alvin nevű katonai tengeralattjáró tanulmányozott, és a hidrotermális kürtőnél burjánzott az élet a nap fénye nélkül világban. Ezeket a kürtőket úgy nevezik, hogy fekete pöfögők. Ezek a pöfögők fekete füstöt okádtak (fémszulfidok) ki magukból. A környezetük erősen savas, valamint hőmérsékletük meghaladja a 400 ºC-ot is. A kürtők kénes ásványi anyagokból, pl. vas-piritből állnak. Innen jött az elnevezés, hogy bolondok aranya, hiszen az anyag színe megegyezik az aranyéval, azonban értékük és keménységük nem igazán. A pöfögőkből felszálló füst anyaga hidrogén-szulfid gáz, amelyből a kürtő mentén burjánzó kénbaktériumok a hidrogént használják fel, amelyet szén-dioxidhoz tudnak kapcsolni, és így hozzák létre a táplálékul szolgáló szerves anyagot. Azonban nemcsak kénbaktériumok tenyésznek itt, hanem egy összefüggő, komplex többsejtű élet, így az oxigén elengedhetetlen, ezért ezt úgy definiálhatjuk, hogy a hidrogén-szulfid és az oxigén reakciója működteti a kürtők világát.
WACHTERHÄUSER (2006) elmélete szerint hidrogén-szulfidból és a vaspiritből származó vas-reakció lehetett az élet legkorábbi felhajtóereje. Azonban a szén-dioxid kevésbé reakcióképes gáz, ezért Wachterhäuser a sokkal reakcióképesebb szén-monoxidot jelölte meg katalizátornak. E módszert laboratóriumban is tesztelték, ahol bámulatos sikereket értek el. Megjelentek az új őslevesben az élet csírái. Azonban kutatók később felfedezték ezen elméletnek is a hibáit. A legnagyobb problémája a szükséges anyagok (vaspirit, hidrogén-szulfid, szén-monoxid) koncentrációs hiányossága okozta, miszerint a csekély számban létrejövő szerves molekulák egy óceánnyi vízben eloszlanának, így nem jöhetett volna létre polimer, sem DNS, sem RNS.
Az elveszett város
Wachterhäuser esetében megismerkedhettünk a hidrtotermális kürtökkel, ezt a módszert próbálta RUSSEL (2004, 2006) is, azzal a különbséggel, hogy az élet eredetét nem a fekete pöfögőkben véli felfedezni, hanem az alkalikus kürtőkben (LANE 2012). A fekete pöfögőkkel ellentétben ezeknek a kürtőknek a környezetében a kémhatás lúgos, a hőmérsékletük meleg, vagy forró, de mégsem 400 ºC.
Russel elmélete szerint az élet alkalikus (lúgos) vegyületeket és nagymértékben redukált hidrotermális oldatokat tartalmazó, egyre növekvő vas-szulfid-buboréktelepekben alakult ki. Erre a legjobb bizonyíték az írországi Tynagh környékéről talált 350 millió éves fosszília a legjobb bizonyíték. Itt a kürtők falát aragonit alkotta. De ugyanilyen környezetet fedeztek fel az Atlanti-óceán közepén húzódó hátságtól mintegy 15 km-re egy víz alatti hegységben, az Atlantis-masszívum nevű helyen. Ezt a helyet a kutatók találóan Elveszett Városnak nevezték el.
Az alkalikus kürtőkben az élet alapja hidrogén és szén-dioxid reakciója volt. A katalizátorok ebben az esetben az oldott vas és kén volt. A hidrogén és szén-dioxid reakciójából ráismerhetünk a fordított (Szent-Györgyi-) Krebs-ciklusnak, amelynek eredeti verziója szerint a ciklus szerves molekulákat (táplálékból) fogyaszt, amelyből hidrogént és szén-dioxidot termel. A fordítottja szerint a kürtőkből feltörő hidrogén csomagok szén-dioxiddal és ATP-vel kiegészülve az élet alapvető építőköveit hozzák létre. Napjainkban a kürtők környezetében élő egyes Archeák még így táplálkoznak. A Krebs-ciklus megfelel a kémia és a termodinamika törvényeinek. Ezzel a ciklussal pedig elindult összetettebb építőanyagok, aminósavak, nukleotidák keletkezése.
Azonban ezen elmélet sem volt teljesen tökéletes, hiszen a hidrogén és a szén-dioxid spontán módon nem reagál egymással. RUSSEL és MARTIN (2007, 2008) pontosították az elvet, miszerint energiabevitelre is szükség van ahhoz, hogy a reakciófolyamat meginduljon. Kutatásaik során felfedezték, hogy a kürtőkből feltörő hidrogén-szulfiddal megjelenő acetil-tioészter a katalizátor. A szén-dioxid spontán módon reagálhat az acetil-tioészterrel, amely energia felszabadulással jár együtt, valamint egy három szénatomos piruvát molekulával. Mivel az alkalikus kürtőkből folyamatos az acetil-tioészter kibocsátása, így kiinduló pont lehetett az élet kialakulásában. Nagy valószínűséggel ez az elmélet áll a legközelebb az élet kialakulásához.
Gánti Tibor (1971) kidolgozott még egy elméletet, a kematon elméletét. E szerint a kematon az élet minimális rendszere, és a legegyszerűbb kémiai alrendszere. Ez a rendszer él, képes alkalmazkodni a környezet változékonyságára, szaporodik és evolúcióképes. Elmélete szerint ezek a rendszerek összehangoltan működnek. Gánti munkáját hazánkban (és külföldön is) jobbára érdektelenség, értetlenség, gúny és rosszindulat fogadta.
Akármelyik formát is vesszük alapul (véleményem szerint az utóbbi, az alkalikus kürtők állnak a legközelebb a valósághoz) kialakult egy közös ős, a Luca (’last universal common ancestor’, utolsó univerzális közös ős), tőle származtatható valamennyi élőlény a Földön (1. ábra).
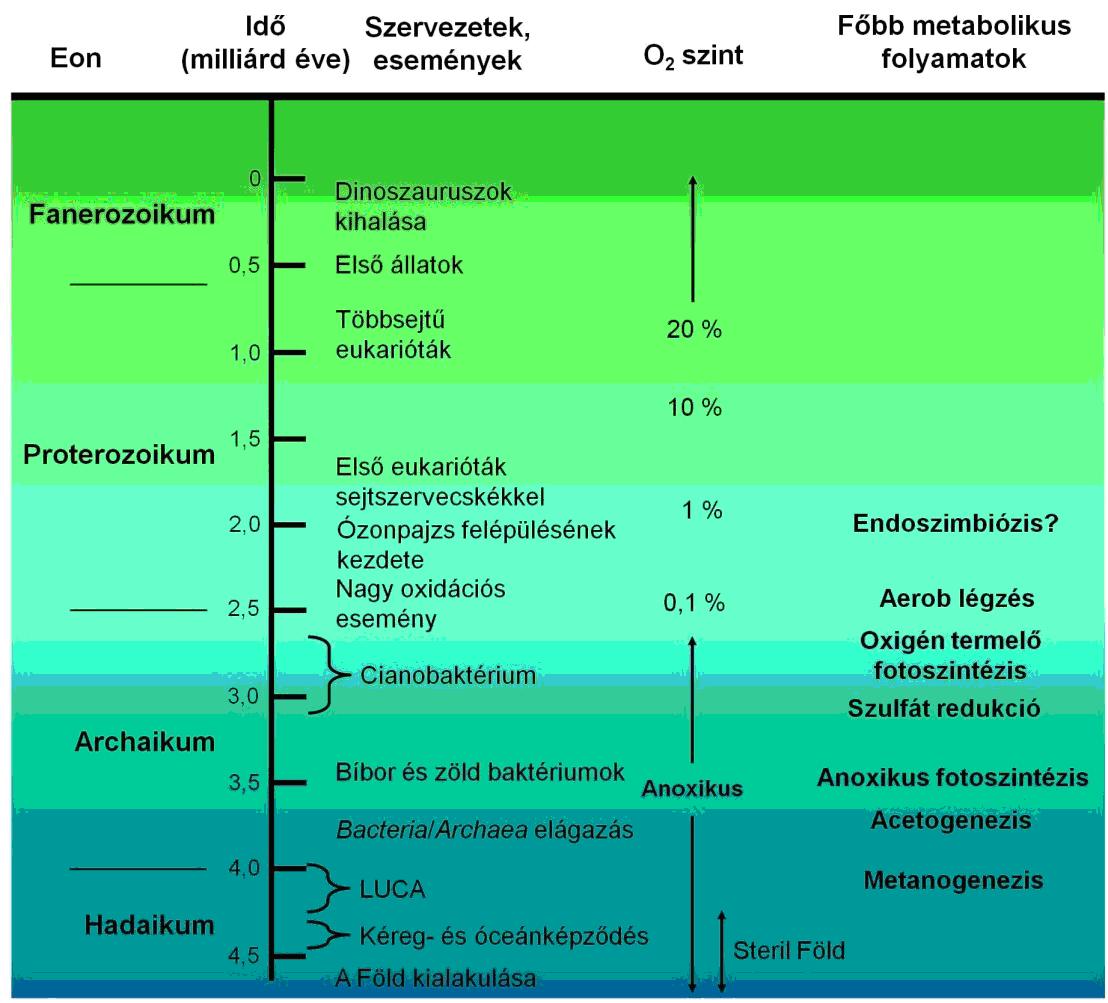 1. ábra A földi élet evolúciójának mérföldkövei (FELFÖLDI T. 2013)
1. ábra A földi élet evolúciójának mérföldkövei (FELFÖLDI T. 2013)
Természetesen az élet csíráinak kialakulása idején még nem voltak komplex szervezetek, majd csak évmilliárdok múlva alakulnak ki a bonyolultabb szervezetek.
A genetikai információtartalom sokszorozódása
Minden ma élő organizmus egyetlen közös ősre vezethető vissza, ez az utolsó közös ős volt, a LUCA. Az egyetlen közös ősből alakult ki a protozoák, az archeoprotozoák és a metazoák. Az evolúció előrehaladása révén létrejöttek az élőlény birodalmak. A négy bemutatott elmélet közül kimagaslik az a nézet, hogy az élet nyilvánvalóan kémiai jelenség, de a különbség nem is a kémiájában rejlik, hanem az informatikai tulajdonságokból ered, hiszen az élő szervezetek komplex információfeldolgozó rendszerek. A komplexitás és az információ a termodinamika tárgyköréhez tartozik, a tudomány azon ágához, amely összekapcsolja a fizikát, a kémiát és a számítástechnikát. Az emberhez képest az egyszerűbb organizmusok kevésbé komplexek, és kevesebb a tennivalójuk, ezért kevesebb genetikus információra van szükségük. Valamely üzenet információtartalmát rendszerint „bit”-nek nevezett egységekben írják le (2. ábra).
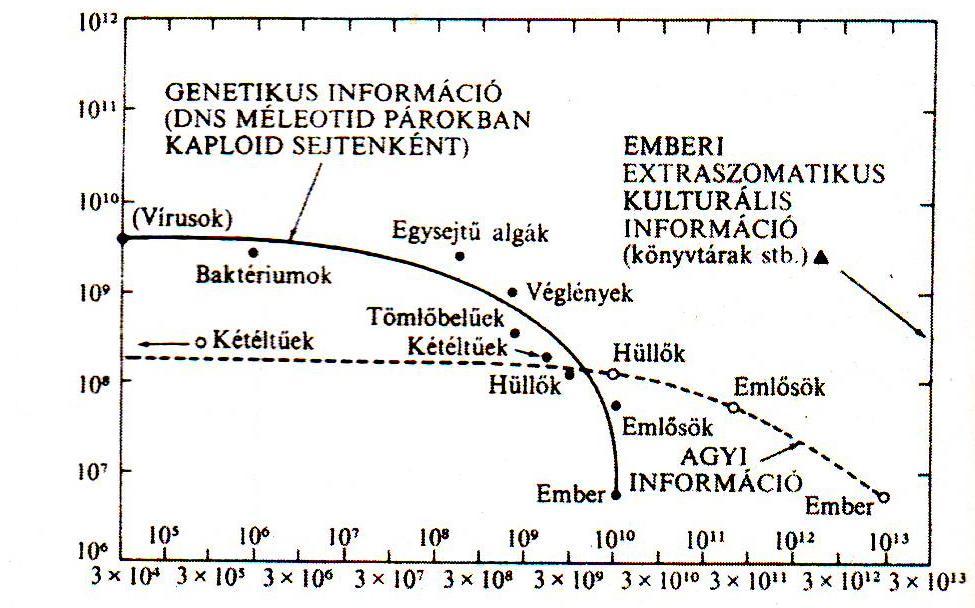 2. ábra A gének és az agyak információtartalmának evolúciója az élet története folyamán (Sagan, C. 2008)
2. ábra A gének és az agyak információtartalmának evolúciója az élet története folyamán (Sagan, C. 2008)
Az ábrából láthatjuk, hogy hárommilliárd évvel ezelőtt meglepően felfokozódott az organizmusok információtartalma, amely fokozatosan nagyon, lassan növekedett tovább. Összességében megállapítható, hogy a molekuláris forgatagot az információ megszervezése jelentette.
Az élet kvantumos alapjai
A következőkben vizsgáljuk meg, hogy a replikáció során, hogyan is tárolódik a csírasejtekben azaz információ, vagy bit, ami a szülőkhöz hasonlóvá teszi az organizmust. E témával Elsasser (1958) foglalkozott behatóan. A központi kérdés az volt, hogy sikerül e a biológiát molekuláris szintre leegyszerűsíteni. Elveti a genetika determinisztikus mivoltát. A biológia és a fizikai ismereteken kívül sokkal inkább hatással vannak rá a filozófiai ismeretek.
A tanulmány nem nyújt kétségbevonhatatlan bizonyosságot. Sok érvet sorakoztat fel amellett, hogy a „lepke, a kígyó, a fa vagy a madár szervezetének felépítése nem vezethető le matematikailag a kromoszómákban tárolt alapvető adatok viszonylag kompakt halmazából” (Wígner 1971). Az információ megőrzése nem írható le megfelelően mechanisztikus következtetésekkel. Elsasser észrevétele szerint az organizmusok összetett rendszerek, amelyeket ha bizonyos tápanyaggal hozunk érintkezésbe, akkor sokszorozódnak, azaz önmagukkal azonos további rendszereket hoznak létre. A vizsgálata arra szorítkozott, hogy hogyan tárolódik a csírasejtekben a felnőtt egyed kifejlődéséhez szükséges információ, és megmutatja, hogy a csírasejtek nem rendelkeznek azokkal a tulajdonságokkal, melyek a fizikus várakozása szerint alkalmassá teszi őket nagy mennyiségű információ tárolására.
Wígner Elsasser tanulmányát is felhasználva megalkotta a fizikusok számára is csoda számba menő eseményt, hogy léteznek olyan rendszerek, amelyek további azonos szerkezettel rendelkező szerkezeteket hoznak létre. Wígner meghatározására azért is volt szükség, mert Elsasser nem fogalmaz meg konkrét javaslatokat a probléma megoldására. Ennek ellenére további természettörvényeket rendel – amelyekből mellesleg teljességgel hiányoznak a fizikai törvények – hozzá, amelyeket biotonikus törvényeknek nevez. Ezek az új törvények nem korlátozzák a kvantummechanika törvényeinek érvényességét. Mivel a kvantummechanika szerint bármilyennek is a viszonyok, mindig alkalmazható és minden értelmes kijelentés belőle vezethető le, így ellentmondás vezethető le.
Ezeket az ellentmondások úgy vezethetők le, hogy a biológiai rendszerek annyira bonyolultak, hogy elvileg lehetetlen kiszámítani viselkedésüket kvantummechanikai törvények alapján. A következőkben nézzük meg az előbbi általános megfontolás numerikus oldalát.
A levezetés nem tökéletesen szigorú, azon alapszik, hogy ha egy rendszer elég bonyolult, akkor nem mutat „véletlen elfajulást”.A számítást két lépésben végezzük el. Először is feltesszük, hogy az élő állapot kvantummechanikailag adott és állapot vektorral leírható állapot. Az állapotvektor komponensét jelölje vx. A tápanyagnak legalább egy olyan állapota kell, hogy létezzen, amely lehetővé teszi az organizmus sokszorozódását. Ezt az állapotot w-vel fogjuk jelölni. A sokszorozódást megelőzően a rendszer, az organizmus + a tápanyag állapotvektora,
φ = v * w, (1)
ahol a kereszt direkt szorzatot jelent. A sokszorozódás után az állapotvektor,
ψ = v * v * r, (2)
két organizmus lesz jelen, mindegyiknek az állapotvektora v, az r vektor egészéről a rendszer azon részét írja le, ami a tápanyagoktól felhasználatlanul hátramarad, s ezen kívül még a két organizmus hely-, stb. koordinátáit. Vezessük be a Hilbert-térben a (2) felbontásának megfelelő koordinátarendszert, ekkor írható:
ψδλη = vδ * vλ * rη, (3)
Az első index (δ) azokat a változókat képviseli, amelyek a rendszerben a „szülő”-ben foglalt részét írják le, a második (λ) a „gyermeket” leíró változók helyettesítője, az utolsó index (η) pedig a rendszer többi részét (hátramaradt tápanyag stb.) leíró változókat helyettesíti. Ugyanebben a koordináta-rendszerben (1) komponensei így írhatók fel:
φδλη = vδ * wλη, (4)
a „tápanyag” állapotát kettős indexekkel jellemezhetjük.
A két feltevés közül az első lényege, hogy a Hilbert-teret véges dimenziós térrel jellemezzük. Az organizmusok N dimenziós térben élnek, a visszamaradott tápanyag pedig R dimenziós. Ebben az esetben az б, λ indexek mindegyike N számú értéket vesz fel, míg az η egyenlő R-rel. Így az organizmusok állapota N dimenziós tér vektora. Az N, R számok nagyságában nincs megszorítás. E felvetés szerint a rendelkezésre álló energia véges, ebből következően a rendszer mindkét része csupán véges számú állapotot érhet el, vagyis az organizmusok tere, egyben a tápanyag R dimenziós tere is (NR).Ebből következően több adatot kell rögzíteni, mint ha csak az organizmusok terét vizsgálnánk. E felvetésből megállapíthatjuk, hogy az élet és az organizmusok sokszorozódása entrópianövekedéssel kapcsolatos, azaz a végállapot kevésbé meghatározott, mint az eredeti állapot. Az organizmus és a tápanyag közötti kölcsönhatás végállapotát az ütközési mátrix jellemzi, amelyet S-sel jelöljük. Minthogy S a (4) alatt megadott φ-t a (3)-mal megadott ψ-be transzformálja:
vбvλrη = Σ Sбλη; б’λ’η’ vб’vλ’η’, (5)
Az S ütközési mátrix az organizmus és a tápanyag közötti kölcsönhatás bármelyik állapotát mutathatja be, így S-t a kvantummechanika törvényei határozzák meg (Elsasser 1958). A gyakorlatban e folyamat úgy néz ki, hogy a б a rendszerben a szülőben foglalt részt jelöli, λ a gyermeket leíró változókat jelöli, η a hátramaradott tápanyagot szemlélteti, egyszóval vбvλrη értéke megadja az ütközési mátrixot, ami szükséges egy újabb organizmus létrejöttéhez. A sokszorozódás állapotvektora (1): φ = v* w, így módosul a φ = v (w1 + w2 + w3). A sokszorozódást követően (2) ψ = v * v * r-re módosul az állapotvektor, hogy két v-let, ahol a v1 a szülő és v2 az utód, míg r a hátramaradott tápanyag. Ha átvezetjük Hilbert-térbe, akkor a (2) számokat б, λ, η helyettesíti. Az (1) képlet koordináta rendszerében is б, λ, η jelöli a változókat.
Elsasser elgondolásai szerint a reprodukciónak az a fajtája, amelyre a Crick és Watson modellje épül, úgy látszik, érvényes, leírható – mint minden hasonló folyamat – az ismert természettörvények segítségével, azonban csak közelítőleg. A modell megbízhatósága a biotonikus törvény következménye, amelyek szuggesztívek, de nem bizonyítható erejűek.
Annak ellenére, hogy Elsasser nem igazolta a biotonikus törvények szükségszerűségét, párhuzamot von Neumann és az – elvileg – véges osztályok között, amely szerint a biológia egyszerűbb folyamatai tele vannak biotonikus jelenségekkel, amelyek nem redukálhatók élesen kvantummechanikai folyamatokkal. Elsasser a körültekintő munkájában egyetlen folyamatot nem vett figyelembe, mégpedig a természetes szelekciót. Elmélete szerint a biológia egy olyan tudomány, amely számítási kapacitást igénylő adatok által vezérelt tudomány.
Davies (2000) tovább lép Elsasser hipotézisén és különbséget tesz a „software” és „hardware” törvények között. Elképzelése szerint a biológiai rendszereket teljes egészében a „software” törvények írják. Az élet alapját két eltérő molekulaosztály között létrejövő megegyezésen alapul. A két molekulaosztály, a nukleinsavak és a fehérjék jelentik. A nukleinsavak tárolják az élet szoftvert, míg a fehérjék, az igazi dolgozók alkotják a hardvert. Így ezek a rendszerek tökéletesen biotonikus tudományok (Gatherer 2008).Összességében elmondhatjuk, hogy Elsasser kvantumszintre csökkentette a mechanizmust. Ha figyelembe vesszük a természetes szelekciót, akkor eldönthetjük, hogy a biológiai folyamatnak vagy determinisztikusnak, esetleg mechaniztikusnak minősül.
Elsasser tanulmányából fontos következtetést vonhatunk le, azonban eredményei nem indokolják a további biotonikus kutatásokat a biológiában, azonban eldönthetetlen, hogy az általa használt fizika (kvantummechanika) helyes-e? A biotonikai kitérő után vizsgáljuk meg, hogy az entrópia milyen módon kapcsolódik az organizmusok születéséhez és halálához.
Az entrópia alakulása
Rudolph Clausius az anyagi rendszerek rendezetlenségére, valamint a termodinamikai mértékre bevezette az entrópia szót. Elsősorban a rendezettség és a rendezetlenség statisztikai összefüggését fejezi ki. Képlettel kifejezve:
S = k log D, (6)
Ahol k a Boltzmann-állandó (1,380 6488(13)•10−23 J/K), és D az anyag egyes részeinek a rendezetlensége (Schrödinger 1987). Köztudott, hogy zárt rendszerben az entrópia törvényszerűen növekszik, míg egy nyílt rendszerben az entrópia alacsonyan tartható úgy, hogy a rendszer saját környezetének hőt ad le. A Földön élő valamennyi élőlény belső egyensúlya az entrópia csökkentésén múlik. Ha az élőlény szervezettségének alacsony állapota felbillen, akkor az oxidációs folyamatokkal már nem képes versenyt folytatni, így entrópiáját nem képes alacsonyan tartani. Amikor az organizmus meghal magas rendezettsége bomlásnak indul, vagyis emelkedik a negatív entrópia miatt. Ha D egyes részek rendezetlenségét jelzi, akkor a reciprok értéke (1/D) egy közvetett rész rendezettségét mutatja. Az elmúlás beállta után bekövetkező negatív entrópiát a következő formában írhatjuk fel:
- (S) = k log (1/D), (7)
Vagyis az entrópia kapcsolatban van a negatív előjellel a rendezettség mértékében is.
Ha egy növény nagy energiájú napfényt vesz fel, és segítségével – CO2 és H2O közreműködésével – cukrokat készít, valamint visszafelé alacsony energiacsomagokban infravörös fényt bocsát ki, akkor az entrópiáját alacsonyan tartja. England (2013) szerint az életfejlődésének kulcsa az evolúciós variáns, miszerint olyan organizmusokra volt szükség, amelyek a jó energiafelhasználásnak köszönhetően képessé váltak az önreprodukcióra, vagy önmásolásra, azaz a szaporodásra. Az RNS-másolása pontosan megfelelt e kritériumoknak. Akár mi is okozta a fajok kialakulását egyetlen közös ősből, az már elmondható, hogy igen komplex életformák alakultak ki a Földön.
A gráfok hálójában
A komplex és bonyolult szervezetek, amelyek az evolúció folyamán kialakultak igen szerteágazóak voltak, azért, hogy rendezhessük őket, hozzárendelhetjük a gráfelméletet, azon belül is a fagráfot. A definíció szerint fáknak nevezzük az összefüggő, kör nélküli gráfokat. Minden gráfnak van egy vagy több csúcsa, de azokat pontosan egy út köti össze. Továbblépve, ha több fa van, akkor egy erdőt képeznek, amelynek bármely két csúcsát egy csak is egy út köti össze. Mivel az erdők komponensei a fák, így az erdők körmentes gráfok. A csúcsokat jelöljük (V), az élek számát pedig E-1-el.
Tételezzük fel, hogy ha G gráf fa, akkor [V] – 1 = [E].E tételnek a bizonyítása pedig így néz ki. A G fa éleinek száma szerinti teljes indukcióval bizonyítunk. Ha a G-nek egy éle van, akkor az állítás igaz. Tételezzük fel, hogy bármely, olyan fára igaz az állítás, melynek legfeljebb n éle van. Tehát érvényes [V1] – 1 = [E1], [V2] – 1 = [E2], és [V3] – 1 = [E3] e három egyenletet összeadva [V1] + [V2] + [V3] – 3 = [E1] + [E2] + [E3] adódik. Figyelembe véve, hogy [V1] + [V2] + [V3] = [V], továbbá [E1] + [E2] + [E3] + 1 = [E].
Próbáljuk meg ezt most egy törzsfejlődést szimbolizáló a 3. ábrán:
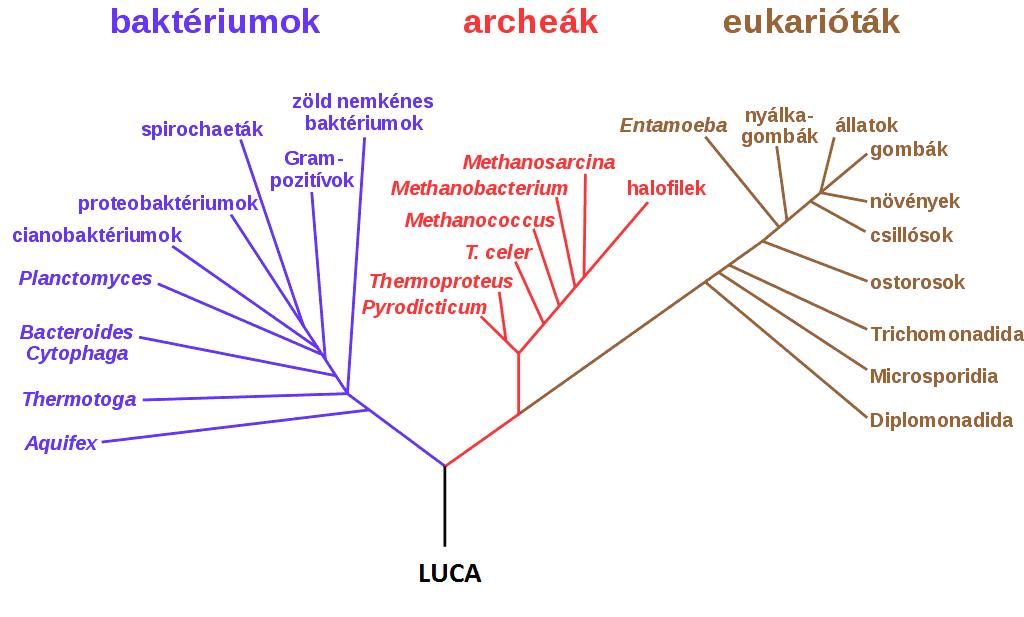 3. ábra Az élővilág törzsfejlődése WOESE és társai (1990) nyomán
3. ábra Az élővilág törzsfejlődése WOESE és társai (1990) nyomán
Sajnálatosan a törzsfejlődést szimbolizáló ábra nem épen a legtökéletesebb, hiszen sokkal több benne az él, mint a csúcs, de vizuális képességünkből adódóan megoldjuk a problémát. Ha megvizsgáljuk a törzsfejlődést szimbolizáló fagráfot, akkor láthatjuk, hogy a fának 27 csúcsa van, amelyből az következik, hogy 24 éle van. Képlettel kifejezve:
[V1] + [V2] + [V3] – 3 = [E1] + [E2] + [E3]; (1)
A LUCA közös ősnek egy foka van. Abban a pontban, ahol az elágazik 3 foka van. A baktériumoknak 9, az archeáknak 7, és az eukariótáknak 10 foka van. A baktériumoknak, az archeáknak és az eukariotáknak a csúcsainak a fokszáma 0, ebből következően izolált pontok.
A következő, a 4. ábrával már sokkal egyszerűbb a helyzet:
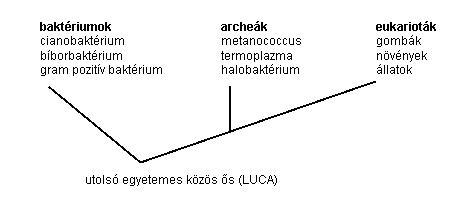 4. ábra Az élővilág fejlődése
4. ábra Az élővilág fejlődése
A 2. ábrán látható, hogy jelen esetben (V =) 4 csúcsunk van és (E = V – 1 =) 3 élünk. Ezen kis ábrán jól áttekinthetjük az élővilág fejlődését egy fagráfon.
A bemutatott módszerrel láthatjuk, hogy a gráfelmélet alkalmazható a biológiában, és az őslénytanban is.
Összefoglalás
Láthatjuk, hogy négy elmélet létezik a földi élet kialakulásáról, és nem is beszélve az extraterresztrikus élet kialakulásának lehetőségéről. Az élet, és a komplexitás kialakulása termodinamika második törvényének engedelmeskedik, miszerint az entrópia fokozatosan növekszik, minél összetettebb organizmusok jönnek létre. Azt azonban vegyük észre, hogy a szerves molekulák sokasága a tengerben olyan lehetett, mint egy növekvő hálózat. Azonban az elmúlt 3 milliárd évben az evolúció és a természetes szelekció érvényesült, ez miatt nem alakulhatott ki skálafüggetlen hálózat, mint a világháló (BARABÁSI A.-L. 2011). Akármilyen bonyolult is lehetett az élet kialakulása a Földön, a fő, hogy kialakult, és fokozatosan fejlődik az evolúció és a természetes szelekció révén. Végső bizonyítékunk azonban még egyik életkeletkezési mechanizmusra sincsen. Ebből következően a nagyszerű osztrák matematikus, Gödel elmélete a legtalálóbb. Talán a legjobb kifejezés az élet kialakulására Gödel IV. nemteljességi tétele, amely így szól:
„Formulák bármely ω-konzisztens rekurzív κ osztályához találhatók olyan r rekurzív osztályok, amelyekben sem a v Gen r, sem pedig Neg (v Gen r) nem tartozik Flg (κ)-hoz (ahol v az r szabad változója).”
Vagyis a matematika bármilyen rendszere, amely véges sok axiómából indul ki, tisztán formális módon épül fel és tartalmaz olyan axiómákat, amely szerint számok sora végtelen: minden számhoz található nála nagyobb szám, és az nem vezethet ellentmondásra, viszont garantáltan tartalmaz olyan állításokat, amelyek a rendszeren belül egzakt módon megfogalmazhatóak, de a rendszeren belül se nem bizonyíthatók, se nem cáfolhatók. Úgy vélem Gödel-tétele pontosan fedezi a valóságot, ami az élet kialakulását illeti, hiszen nem láttuk, hogy az élet milyen módon is alakult ki, csupán implicit bizonyítékunk van róla.
Végső soron a kialakuló új paradigma, holisztikusan szemléli a világot, mivel a világ egy egységes egész, nem pedig egymástól elszigetelten, egymástól elkülönült részek egésze.
Irodalom
- BARABÁSI A.-L. 2011: Behálózva. A hálózatok új tudománya. – Helikon Kiadó, 201-205.
- CAIRNS-SMITH, G. 1982: Genetic Takeover. – Cambridge: Cambridge University Press.
- CAIRNS-SMITH, G. 1985: Seven Clues to the Origin of Life. – Cambridge: Cambridge University Press.
- DAVIES, P. 2000: Az ötödik csoda. Vince Kiadó. 11-34.
- DAWKINS, R. 2011: A vak órásmester. – Kossuth Kiadó, 195-219.
- ELSASSER, W. M. 1958: The Physical Foundations of Biology. An Analytical Study. London: Pergamon Press.
- ENGLAND, J. 2013: Statistical physics of self-replication. – The Journal of Chemical Physics, 139. 121923-2, 1-9.
- FELFÖLDI T. 2013: A mikroorganizmusok világa és a bioszféra koevolúciója. – In: MÁRIALIGETI K. (szerk.) 2013: Bevezetés a prokarióták világába. – Eötvös Lóránd Tudományegyetem, Budapest. http://elte.prompt.hu/sites/default/files/tananyagok/prokariotak/index.html
- GATHERER, D. 2008: Finite Universe of Discourse: The Systems Biology of Walter Elsasser (1904- 1991). The open Biology Journal, 1, 9-20.
- GÁNTI T. 1971: Az élet princípiuma. – Gondolat, Budapest.
- HOYLE, F. 1972: A fekete felhő. – Gondolat Kiadó, Budapest.
- MARTIN, W. – BAROSS, J. – KELLEY, D. – RUSSEL, M. J. 2008: Hydrothermal vents and the origin of life. – Nature Reviews in Microbiology, 6, 805-814.
- MARTIN, W. – RUSSEL, M. J. 2007: Ont he origin of biochemistry at an alkaline hydrothermal vent. – Philosopical Transaction of the Royal Society of London, B 362, 1887-1925.
- UREY, H. – MILLER, S. L. 1953: Production of Amino Acids Under Possible Primitive Earth Conditions. Science, 117, p. 528.
- SAGAN, C. 2008: Az Éden sárkányai. Európa Kiadó, 55-60.
- SCHRÖDINGER, E. 1987: Was ist leben? – R. Piper GmbH & Co. KG, München, 126 p.
- VON NEUMANN J. 1961: Uber die Grundlagen der Quantenmechanik. In: Taub AH, Ed: John von Neumann Collected Works Vol 1: Logic, Theory of Sets and Quantum Mechanics. Oxford Pergamon Press.
- WÄCHTERHÄUSER, G. 2006: From volcanic origins of chemoautotrophic life to bacteria, archea and eukarya. – Philosopical Transaction of the Royal Society of London, B 361, 1787-1806.
- WIGNER J. 1971: Szimmetriák és reflaxiók. Gondolat Kiadó.
- WOESE, C. R., KANDLER, O., WHEELIS, M. L. 1990: Towards a natural system of organisms: proposal for the domain Archeae, Bacteria and Eukarya. – Proceedings of the National Academy of Sciences of the United States of America 87 (12), 4576–4579










Az utolsó 100 komment: